Ongoing GAN experiments with Marco Pisano.
Last update 30.03.23.
reducing the file size of an h264 video in ffmpeg.
ffmpeg -i input.mp4 -vcodec libx264 -crf 27 -preset slower output_reduced_in_size.mp4













hex code for silver is: #C0C0C0
from: https://people.xiph.org/~xiphmont/demo/neil-young.html
Unfortunately, there is no point to distributing music in 24-bit/192kHz format.
Its playback fidelity is slightly inferior to 16-bit /44.1 or 16/48, and it takes up 6 times the space.
Ears and auditory concentration both fatigue quickly, so testing must be done when a listener is fresh. That means lots of breaks and pauses. Testing takes anywhere from many hours to many days depending on the methodology.
The upper limit of the human audio range is defined to be where the absolute threshold of hearing curve crosses the threshold of pain. To even faintly perceive the audio at that point (or beyond), it must simultaneously be unbearably loud.
At low frequencies, the cochlea works like a bass reflex cabinet. The helicotrema is an opening at the apex of the basilar membrane that acts as a port tuned to somewhere between 40Hz to 65Hz depending on the individual. Response rolls off steeply below this frequency.
The visible spectrum extends from about 400THz (deep red) to 850THz (deep violet)
All signals with content entirely below the Nyquist frequency (half the sampling rate) are captured perfectly and completely by sampling; an infinite sampling rate is not required. Sampling doesn't affect frequency response or phase. The analog signal can be reconstructed losslessly, smoothly, and with the exact timing of the original analog signal.


First week of April '22, experimenting with led wall & touchdesigner.
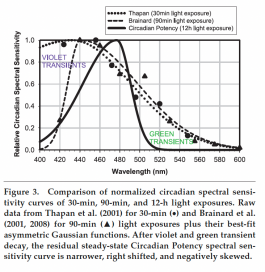
from: "Circadian Potency Spectrum with Extended Exposure to Polychromatic White LED Light under Workplace Conditions" - Martin Moore-Ede, Anneke Heitmann and Rainer Guttkuhn.
With mounting evidence that artificial light sources can disrupt circadian rhythms and lead to myriad negative health outcomes, there is a dire need for biologically healthy light sources (Zielinska-Dabkowska, 2018). There are several problems with using the previously reported circadian spectral sensitivity curve from short exposures of dark-adapted subjects to monochromatic light to spectrally engineer healthy white light sources for day and night applications.
First, the previously reported circadian spectral sensitivity from dark-adapted subjects spans a broad band of violet, blue, and green wavelengths (400-560nm), and when all of these wavelengths are removed from the polychromatic 380- to 780-nm white light spectrum, the result is an unattractive yellow-orange light not suitable for most human uses.
The definition of the steady-state Circadian Potency spectral sensitivity curve makes it feasible to design and build blue-rich white LED chips to optimize Circadian Potency during daytime hours and bluedepleted white LED chips to minimize Circadian Potency from sunset to sunrise. Because the steadystate Circadian Potency spectral sensitivity is narrower than the previously reported circadian spectral sensitivity curves for dark-adapted conditions, it permits the use of dies and phosphors that emit violet and green (plus yellow and red) wavelengths at night to produce attractive and energy-efficient white electric light, which minimizes circadian disruption.
One particularly interesting feature of the Circadian Potency spectral sensitivity curve is the relative insensitivity to spectral wavelengths less than 420 nm. This permits the incorporation of 2 important features into nocturnal circadian lighting. First, violet LED dies with peak wavelengths of 410 to 420 nm can be used to replace the typical ~450-nm blue peak emissions of conventional LEDs to spectrally engineer colorimetrically balanced attractive white-colored light.
Second, short-wavelength light is known to have alerting-, performance-, and moodenhancing properties (Rahman et al., 2014; Viola et al., 2008), leading to the proposal that blue-rich light should be used at night to reduce human error, but this has the unfortunate consequence that it could significantly increase the risk of circadian disruption and health disorders.
There is evidence that the alerting effects of 420-nm violet light are even greater than 440-nm or 470-nm blue light (Revell et al., 2006).
Why might evolutionary selection result in such a narrow band of blue wavelengths defined by the Circadian Potency spectral sensitivity curve being used to communicate the environmental day-night signal? We suggest it may be because life forms have been exposed to steady-state blue wavelengths during the day ever since the origin of life deep in the primitive oceans. Below a depth of 200 m, all other visible spectral wavelengths except for ~475-nm light are absorbed by seawater (Denton, 1990). From single- cell marine organisms such as Gonyaulax (Hastings, 2001) to humans, the fundamental apparatus consisting of (1) blue photoreceptors maximally sensitivity at ~475 nm, (2) circadian clocks, and (3) melatonin release as the internal temporal signal has been conserved for billions of years to equip life for successful predictive adaptation to a rotating planet (Moore-Ede, 1986).